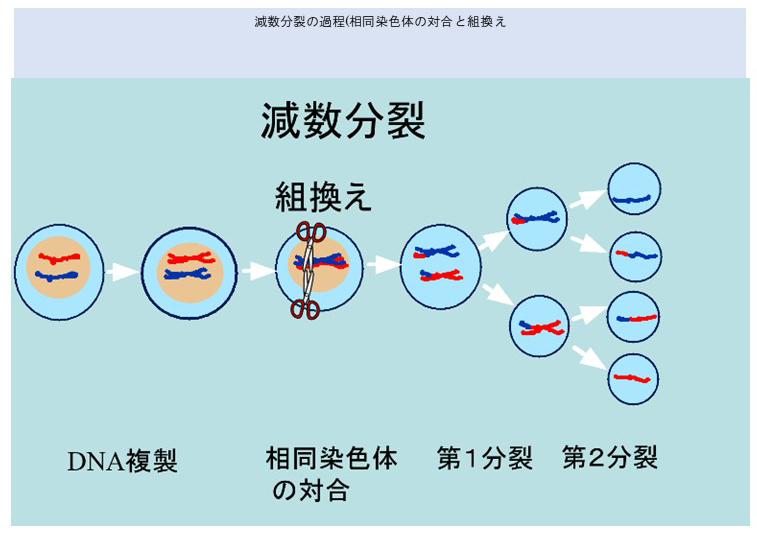
減数分裂
遺伝子の組換えは、本来生殖細胞に特有な現象と考えられ、卵子や精子などの配偶子を形成するときにおこる減数分裂に重要な過程である。
減数分裂では、父由来と母由来の染色体が対合を起こし、その後キアズマという場所で組換えがおこる。
キアズマは染色体の接着部位であり、その後の染色体分離に欠かせず、組換えは減数分裂にとって必須な出来事といえる。
キアズマは染色体あたり1から2,3か所である。
この組換えにより配偶子の遺伝子の多様性は増加するため、有性生殖の重要な役割を担っているといえる。
下図で、減数分裂の過程を示す。
このような減数分裂における組換え遺伝子として、先に述べたRad51遺伝子は重要であるが、その他に、Dmc1とよばれるRAD51タンパクと非常に構造の似たタンパクをコードする遺伝子が哺乳類でもクローニングされた。
その発現は精子形成の場合、減数分裂前期のレプトテン期という時期に発現していた。
我々はそのDmc1遺伝子のノックアウトマウスを作製した。
その結果、精巣の切片の写真のように、Dmc1遺伝子ノックアウトマウスは、レプトテン期で減数分裂を停止しアポトーシスを起こすことがわかった。
さらに電子顕微鏡による染色体解析の結果、父母染色体の対合がみられないことから、減数分裂特異的相同組換え遺伝子Dmc1は減数分裂期に起こる染色体対合に必要であることを明らかにした。
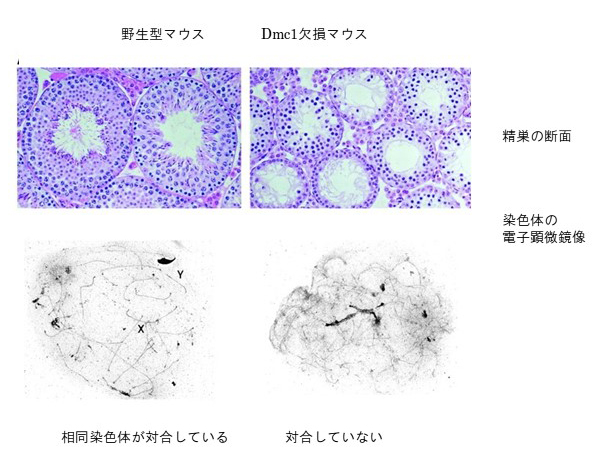
文献;The mouse RecA-like gene Dmc1 is required for homologous chromosome synapsis during meiosis.
Yoshida K, Kondoh G, Matsuda Y, Habu T, Nishimune Y, Morita T.: Mol Cell. 1998 1:707-18.
さらに、RAD51とDMC1抗体を用いた実験から、減数分裂期には、RAD51とDMC1タンパクが相同染色体の対合部位である組換え小節という部分にあることが明らかになった。
減数分裂が進むと組換え小節のいくつかがキアズマになるといわれているので、キアズマにおいてRAD51とDMC1両タンパクが染色体組換えをしていると考えられる。
両方のタンパクは構造が似ていることから、DMC1もRAD511と同様なメカニズムで組換えを起こしている可能があるので我々は次のようなモデルを考えている。
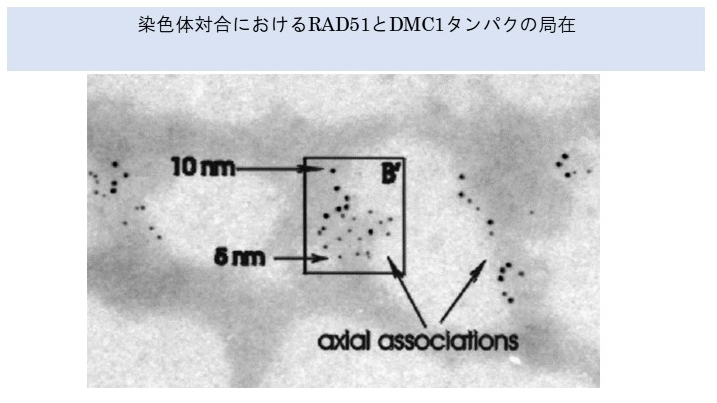
文献;RAD51 and DMC1 form mixed complexes associated with mouse meiotic chromosome cores and
Yoshida K, Kondoh G, Matsuda Y, Habu T, Nishimune Y, Morita T.: Mol Cell. 1998 1:707-18.
(結語) このように、相同組換え(修復)遺伝子は、Rad51のように細胞分裂と連携してDNA二重鎖切断を修復し、遺伝情報の恒常性のために貢献している。
一方、減数分裂特異的なDmc1遺伝子は配偶子形成において染色体の二重鎖切断(Spo11による)を修復する過程において父母染色体間に組換えを起こさせ、配偶子の遺伝子の多様性をもたらすという恒常性とは全く逆の機能をもたらすと考えられ興味深い。